为什么研究snoRNAs?
产品名称: 为什么研究snoRNAs?
英文名称: snoRNAs
产品编号:
产品价格: 0
产品产地: null
品牌商标: null
更新时间: null
使用范围: null
- 联系人 :
- 地址 : 上海市宜山路700号C2栋3楼 (200233)
- 邮编 : 200233
- 所在区域 : 上海
- 电话 : 00 点击查看
- 传真 : 点击查看
- 邮箱 : wangyanling@kangchen.com.cn;market@kangchen.com.cn
引言
核仁小RNA(snoRNAs)是一类中等长度的非编码小RNA,它们的长度在60-300nt不等,能与核仁核糖核蛋白结合形成snoRNPs 复合物[1]。在脊椎动物中编码核仁小RNA的基因主要存在于蛋白编码基因或非蛋白编码基因的内含子区域,并且经过进一步的转录后加工处理形成成熟的核仁小RNA[2]。snoRNAs参与的生物学过程主要有rRNA的加工处理,RNA剪接和翻译过程的调控以及氧化应激反应[3]。近期的研究表明snoRNAs还参与到遗传性疾病[4]、人类的变异[5]、造血[6]、代谢[7, 8]以及癌症[3, 9]的过程中。
snoRNAs的生物合成
snoRNAs主要包含两个家族:C/D box snoRNAs 与H/ACA box snoRNAs (Fig.1A and Fig.1B)。大多数snoRNAs主要位于由RNA聚合酶II转录的基因的内含子区域(Fig.1C)[9]。但snoRNAs也可以来源于长链非编码RNA(lncRNA)的内含子区域。例如GAS5 编码9个不同的C/D box snoRNAs(snoRNDs74-81)[10]。从内含子上脱离后,pre-snoRNAs进一步的被核酸外切酶处理去除两端的多余序列,进而形成成熟的snoRNAs.。snoRNAs内部的信号序列指导snoRNAs与相应蛋白结合形成snoRNP复合物来避免被酶切,进而发挥功能。
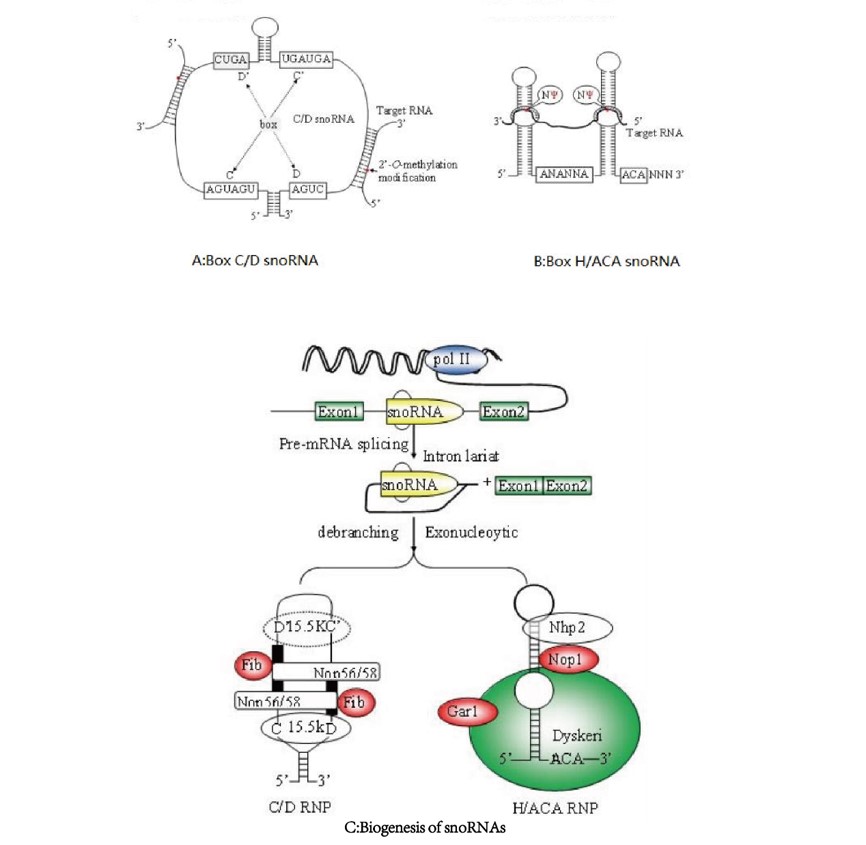
Fig1: The structure and biogenesis of snoRNAs. A. The 2′-O-methylated nucleotides located five nucleotides upstream of the D or D’ box sequences are indicated as red. B. Positions and consensus sequences of the conserved C, C’ box (UGAUGA), and D, D’ boxes (CUGA). The uridine residues selected for pseudouridylation are shown as. C. Biogenesis of snoRNAs :The great majority of mammalian box C/D and H/ACA snoRNAs are processed from pre-mRNA introns. snoRNAs families are not independently transcribed but processed from the pre-mRNA introns, in most cases by exonucleolytic digestion of the debranched lariat. Box C/D snoRNAs contain four evolutionarily conserved, essential proteins, fibrillarin (methyltransferase), Nop56, Nop58, and 15.5kDa. Proteins common to H/ACA snoRNAs include dyskerin (pseudouridine synthase), Gar1, Nhp2, and Nop10p[9].
snoRNAs的功能
snoRNAs在核糖体RNA加工过程的作用
在核糖体RNA的加工过程中,snoRNAs发挥这两个基本的作用:核糖体RNA的2′ -O –甲基化和假尿嘧啶化[11]。核糖体RNA的2′ -O –甲基化由C/D box snoRNAs来负责。 C/D box snoRNAs 包含有两个短的序列元件,即位于5 " 末端的box C(RUGAUGA R代表A或G)和3" 末端的box D(CUGA)。多数box C/ D snoRNAs基因的5" 和3" 末端都有4 - 5 nt 的反向重复序列,可以形成较为稳定的短茎结构,这一结构在snoRNAs的生物合成和核仁定位过程中起关键作用(Fig. 1A)[12]。大部分C/D box snoRNAs的中部,还具有类似于box C 和box D 的结构,通常分别表现出 1 - 2 nt的差异,被称为box C" 和box D"[13]。核糖体RNA的假尿嘧啶修饰主要是由H/ACA box snoRNAs来完成的。H/ACA box snoRNAs 具有保守的“发夹- 铰链- 发夹- 尾(hairpin-hinge hairpin -tail ) ”的二级结构( Fig.1B)[13] 。box H (ANANNA,N 代表任一核苷酸)位于单链形式的铰链区,ACA 则一般位于3" 末端上游3个核苷酸处。H/ACA box snoRNAs的指导序列位于单个或者两个发夹内部的“假尿嘧啶化泡”(pseudouridylation pocket) 内,它们可以与靶RNA上的被修饰位点两翼序列各形成4 - 8nt 的互补配对。另外还有一种比较特殊的snoRNAs- SCARNAs,只特异存在于Cajal小体中,能够对剪接体RNA进行2′ -O –甲基化和假尿嘧啶化修饰[14]。
snoRNAs作为miRNA前体
越来越多的实验结果表明snoRNAs可以进一步被加工形成更短片段的RNA,并且这些短片段的snoRNAs具有类似miRNA的功能,这就表明snoRNAs可能作为miRNA的前体[15]。ACA45是第一个被报道能够被降解成短片段的snoRNAs,它是通过与Ago蛋白的相互作用被发现的,而Ago作为miRNA RISC复合物的成员,这就表明ACA45的进一步的加工处理是依赖于Dicer酶的。进一步的实验表明,降解产生的20-22nt的短片段RNA的作用机制类似与miRNA-抑制目的基因(CDC2L6)的表达[15]。近期报道发现11个C/D box snoRNAs能够降解形成短片段RNA从而抑制靶基因的表达[16]。
snoRNAs在可变剪切过程中的作用
snoRNAs既能作为miRNA的前体也可以调节RNA的可变性剪切。研究报道HBII-52 C/D box snoRNAs上的18个核苷酸能够与5-羟色胺受体5-HT (2C)的mRNA序列互补配对进而通过影响5-羟色胺受体5-HT (2C)的可变剪切来调控5-羟色胺受体的表达,最终导致Prader-Willi综合征[17]。
snoRNAs在应急反应过程中的作用
核仁是细胞应对应急反应的主要参与者。不同的压力条件能够导致核仁的变化甚至破坏它。据报道,snoRNAs有助于细胞应对应急反应。在缺氧条件下,SNORD14A 和SNORD83B的表达水平得到显著提高[18]。另有报道发现,棕榈酸酯处理能够导致细胞中SNORDs 32A, 33, 和35A的表达水平也显著提高。细胞对棕榈酸酯产生抗性是由于抑制了SNORDs 32A, 33, 和35A的表达,它们介导了由诱导剂引起的细胞死亡[7]。只有在细胞应急反应的早期过程中能够在细胞质中检测到snoRNAs的存在。例如,在代谢压力的早期过程中,相互独立的snoRNAs(SNORD3, SNORD13和 SNORD118)以及它们各自的靶向mRNA在细胞质中相互依存并且调节它们的翻译。snoRNAs参与调节的细胞过程将会更加复杂。
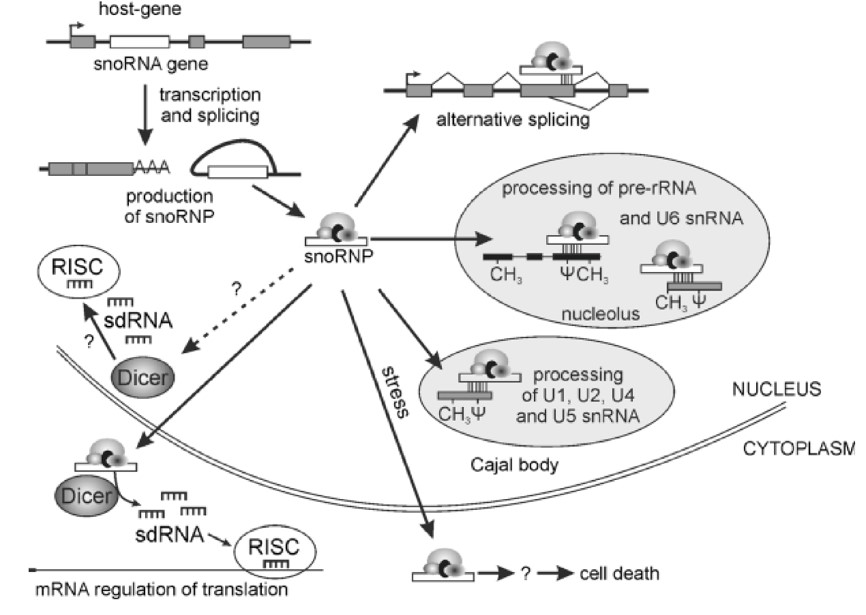
Fig2: snoRNAs参与的细胞过程[19]
snoRNAs在癌症中的作用
snoRNAs参与的癌症的分子病理学
研究snoRNAs在癌症中的作用起始于一项研究:在脑膜瘤中,与正常脑组织相比snoRNAs的表达大幅下调[20]。最近研究发现,在非小细胞肺癌中多种snoRNAs呈现出不同的表达状态[21]。其它的研究证明snoRNAs U50 2bp纯合缺失突变与前列腺癌的发生发展有关[22]并且在乳腺癌中U50具有杂合缺失以及转录下调的特点[23]。另有研究发现snoRNAs以及lncRNA的hostgene-GAS5能够调控细胞的生存通过诱导或者感受细胞的凋亡。GAS-5在乳腺癌中大幅下调的暗示其可能作为抑癌基因存在[24]。snoRNAs与肿瘤的关系也能够延伸到与它们相互作用的蛋白分子上。根据它们结构与功能的不同,snoRNP复合物能够分为两类C/D box snoRNP和H/ACA box snoRNP。C/D box snoRNP 包含四种进化上保守的,必需的蛋白质,即纤维蛋白 (fibrillarin)/Nop1p、Nop56、Nop58/Nop5p和p15.5KD/ Snu13p,而H/ACA box snoRNP 的结合蛋白包括进化保守的Cbf5p(dyskerin)、Gar1p、Nhp2p 和Nop10p。fibrillarin是发育必须的,缺失fibrillarin能够导致胚胎致死。Dyskerin、NOP10以及Nhp2p的突变与上皮癌有关[25]。snoRNAs以及snoRNP很可能是通过影响核糖体和蛋白的翻译来影响肿瘤的发生的,因为在癌细胞中翻译过程总是异常的。但是snoRNAs也可能通过降解的短片段RNA来发挥miRNA功能进而调节基因的表达来影响肿瘤的发生。孤儿snoRNAs(目前还没有明确的靶基因)可能成为以后研究的热点。
snoRNAs作为癌症诊断和预后生物标志物
snoRNAs在细胞中的功能是多样的,肿瘤特异表达的snoRNAs可能作为癌症相关的生物标志物。研究报道snoRNAs能够在外周血浆以及血清中稳定存在且可测,这使snoRNAs作为潜在的循环肿瘤生物标志物成为可能(table1)[9]。在非小细胞肺癌中 SNORD33, SNORD66和SNORD76不仅在肿瘤中高表达并且在血浆中也检测到高表达,这使辨别非小细胞肺癌患者成为可能[21]。SNORA42虽在组织相关疾病的预后检测效果较差,但可以预测疾病[26]。近期的研究成果进一步表明snoRNAs作为生物标志物的巨大应用前途。他们发现SNORD43, SNORD44和SNORD48作为乳腺癌和头颈部鳞状细胞癌潜在的抑制剂[27]。U50能够抑制乳腺癌和前列腺癌(table1)[22, 23]。SNORD113-1在原发性肝细胞癌中表达下调并且在原发性肝细胞癌中作为抑癌基因发挥功能[28]。总之,snoRNAs可以作为癌症诊断和预后的潜在的生物标志物。
snoRNAs作为癌症治疗的潜在靶点
虽然snoRNAs在肿瘤中发挥功能的分子机制仍然不是很清楚,但其仍是作为基因干预治疗的潜在理想对象。由snoRNAs介导的基因沉默可能是最好的治疗手段。当然用RNAi介导的基因沉默来选择性的沉默相关致癌snoRNAs也是不错的方法。例如,snoRA42在肺癌中是高表达的,在肺癌细胞中选择性的沉默snoRNAs42,细胞的增殖能力和活力明显下降[26]。这些实验证据都表明snoRNAs可以作为癌症治疗的潜在靶点。
snoRNAs和其它疾病
snoRNAs与神经退行性疾病的关系也有报道[29, 30]。一系列独立的研究表明Prader Willi综合症是由于缺失来源于15号染色体15q11–q13区域所致,而此区域则包含两个snoRNAs家族成员:SNORD115 (HBII-52) 和SNORD116 (HBII-85)[31]。SNORD115可能影响大脑中5-HT2CR的mRNA水平[17]。SNORD116的缺失是导致Prader Willi综合症发病的主要原因。最近的一项研究表明,C/D box snoRNAs 表达水平的变化也发生在孕妇酒精消耗引起的异常胎儿大脑发育的过程中。特别注意的是在此过程中SNORD115表达上调,SNORD116表达下调[32]。SNORD115表达水平的变化导致了各种心理和行为畸变典型自闭症[33]。同时另外一项研究报道在病毒感染的细胞中一系列snoRNAs的表达水平在病毒感染后得到明显上调[34]。一方面snoRNAs可以作为受体抗病毒反应的调节者;另一方面,调控RNA的活性可以被病毒利用使其逃避天然免疫从而完成其生命周期[35]。
总结
在人类细胞中的,RNA的转录后修饰、mRNA的稳定性和翻译的控制是基因表达调控的重要组成部分。snoRNAs和它们产生的功能短片段RNA在这些过程中发挥着重要作用:它们指导核糖体RNA(rRNA)和核小RNA(snRNA)的修饰、影响其互补前体mRNA的剪接以及控制mRNA的翻译和稳定性。外界因素以及细胞内部的信号级联反应能够导致snoRNAs表达水平的变化,进而引起细胞水平的生理变化、器官功能障碍以及各种疾病。snoRNAs的结构,表达模式和在细胞中的定位具有重要的调控作用,被认为是病理诊断的标志物。snoRNAs表达水平和功能机制的研究将为人类疾病的诊断和治疗提供新的希望。
参考文献
1. Esteller, M., Non-coding RNAs in human disease. Nat Rev Genet, 2011. 12(12): p. 861-74.
2. Kiss, T., et al., Biogenesis and intranuclear trafficking of human box C/D and H/ACA RNPs. Cold Spring Harb Symp Quant Biol, 2006. 71: p. 407-17.
3. Williams, G.T. and F. Farzaneh, Are snoRNAss and snoRNAs host genes new players in cancer? Nat Rev Cancer, 2012. 12(2): p. 84-8.
4. Sahoo, T., et al., Prader-Willi phenotype caused by paternal deficiency for the HBII-85 C/D box small nucleolar RNA cluster. Nat Genet, 2008. 40(6): p. 719-21.
5. Bhartiya, D., et al., Systematic curation and analysis of genomic variations and their potential functional consequences in snoRNAs loci. Hum Mutat, 2012. 33(10): p. E2367-74.
6. Bellodi, C., et al., H/ACA small RNA dysfunctions in disease reveal key roles for noncoding RNA modifications in hematopoietic stem cell differentiation. Cell Rep, 2013. 3(5): p. 1493-502.
7. Michel, C.I., et al., Small nucleolar RNAs U32a, U33, and U35a are critical mediators of metabolic stress. Cell Metab, 2011. 14(1): p. 33-44.
8. Youssef, O.A., et al., Potential role for snoRNAss in PKR activation during metabolic stress. Proc Natl Acad Sci U S A, 2015. 112(16): p. 5023-8.
9. Mannoor, K., J. Liao, and F. Jiang, Small nucleolar RNAs in cancer. Biochim Biophys Acta, 2012. 1826(1): p. 121-8.
10. Smith, C.M. and J.A. Steitz, Classification of gas5 as a multi-small-nucleolar-RNA (snoRNAs) host gene and a member of the 5'-terminal oligopyrimidine gene family reveals common features of snoRNAs host genes. Mol Cell Biol, 1998. 18(12): p. 6897-909.
11. Panse, V.G. and A.W. Johnson, Maturation of eukaryotic ribosomes: acquisition of functionality. Trends Biochem Sci, 2010. 35(5): p. 260-6.
12. Thorenoor, N. and O. Slaby, Small nucleolar RNAs functioning and potential roles in cancer. Tumour Biol, 2015. 36(1): p. 41-53.
13. Kiss, T., Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs. EMBO J, 2001. 20(14): p. 3617-22.
14. Ganot, P., M.L. Bortolin, and T. Kiss, Site-specific pseudouridine formation in preribosomal RNA is guided by small nucleolar RNAs. Cell, 1997. 89(5): p. 799-809.
15. Ender, C., et al., A human snoRNAs with microRNA-like functions. Mol Cell, 2008. 32(4): p. 519-28.
16. Brameier, M., et al., Human C/D box snoRNAss with miRNA like functions: expanding the range of regulatory RNAs. Nucleic Acids Res, 2011. 39(2): p. 675-86.
17. Kishore, S. and S. Stamm, The snoRNAs HBII-52 regulates alternative splicing of the serotonin receptor 2C. Science, 2006. 311(5758): p. 230-2.
18. Liu, Z.H., et al., Small ncRNA expression and regulation under hypoxia in neural progenitor cells. Cell Mol Neurobiol, 2011. 31(1): p. 1-5.
19. Makarova, J.A., et al., New functions of small nucleolar RNAs. Biochemistry (Mosc), 2013. 78(6): p. 638-50.
20. Chang, L.S., et al., Differential expression of human 5S snoRNAs genes. Biochem Biophys Res Commun, 2002. 299(2): p. 196-200.
21. Liao, J., et al., Small nucleolar RNA signatures as biomarkers for non-small-cell lung cancer. Mol Cancer, 2010. 9: p. 198.
22. Dong, X.Y., et al., Implication of snoRNAs U50 in human breast cancer. J Genet Genomics, 2009. 36(8): p. 447-54.
23. Dong, X.Y., et al., SnoRNAs U50 is a candidate tumor-suppressor gene at 6q14.3 with a mutation associated with clinically significant prostate cancer. Hum Mol Genet, 2008. 17(7): p. 1031-42.
24. Mourtada-Maarabouni, M., et al., Growth arrest in human T-cells is controlled by the non-coding RNA growth-arrest-specific transcript 5 (GAS5). J Cell Sci, 2008. 121(Pt 7): p. 939-46.
25. Gupta, V. and A. Kumar, Dyskeratosis congenita. Adv Exp Med Biol, 2010. 685: p. 215-9.
26. Mei, Y.P., et al., Small nucleolar RNA 42 acts as an oncogene in lung tumorigenesis. Oncogene, 2012. 31(22): p. 2794-804.
27. Gee, H.E., et al., The small-nucleolar RNAs commonly used for microRNA normalisation correlate with tumour pathology and prognosis. Br J Cancer, 2011. 104(7): p. 1168-77.
28. Xu, G., et al., Small nucleolar RNA 113-1 suppresses tumorigenesis in hepatocellular carcinoma. Mol Cancer, 2014. 13: p. 216.
29. Doe, C.M., et al., Loss of the imprinted snoRNAs mbii-52 leads to increased 5htr2c pre-RNA editing and altered 5HT2CR-mediated behaviour. Hum Mol Genet, 2009. 18(12): p. 2140-8.
30. Sridhar, P., H.H. Gan, and T. Schlick, A computational screen for C/D box snoRNAss in the human genomic region associated with Prader-Willi and Angelman syndromes. J Biomed Sci, 2008. 15(6): p. 697-705.
31. Cavaille, J., et al., Identification of brain-specific and imprinted small nucleolar RNA genes exhibiting an unusual genomic organization. Proc Natl Acad Sci U S A, 2000. 97(26): p. 14311-6.
32. Laufer, B.I., et al., Long-lasting alterations to DNA methylation and ncRNAs could underlie the effects of fetal alcohol exposure in mice. Dis Model Mech, 2013. 6(4): p. 977-92.
33. Nakatani, J., et al., Abnormal behavior in a chromosome-engineered mouse model for human 15q11-13 duplication seen in autism. Cell, 2009. 137(7): p. 1235-46.
34. Saxena, T., et al., Combined miRNA and mRNA signature identifies key molecular players and pathways involved in chikungunya virus infection in human cells. PLoS One, 2013. 8(11): p. e79886.
35. Cullen, B.R., MicroRNAs as mediators of viral evasion of the immune system. Nat Immunol, 2013. 14(3): p. 205-10.
地址:上海市漕河泾高新技术开发区虹漕路421号63号楼2楼
免费热线:400-886-5058 ; 800-820-5058(座机)
电话:021-64451989 传真:021-64452021